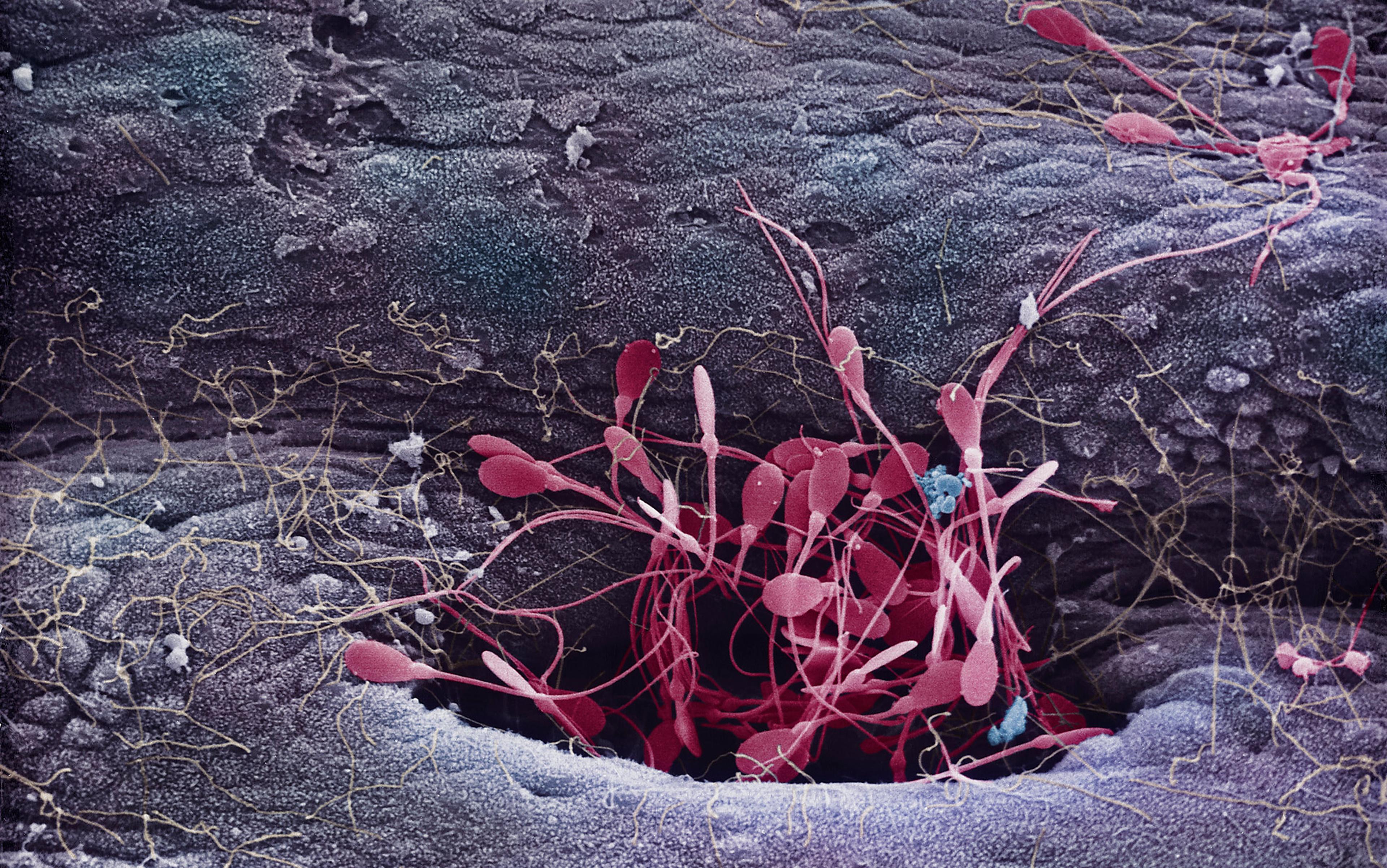
It’s uncontroversial among biologists that many species have two, distinct biological sexes. They’re distinguished by the way that they package their DNA into ‘gametes’, the sex cells that merge to make a new organism. Males produce small gametes, and females produce large gametes. Male and female gametes are very different in structure, as well as in size. This is familiar from human sperm and eggs, and the same is true in worms, flies, fish, molluscs, trees, grasses and so forth.
Different species, though, manifest the two sexes in different ways. The nematode worm Caenorhabditis elegans, a common laboratory organism, has two forms – not male and female, but male and hermaphrodite. Hermaphroditic individuals are male as larvae, when they make and store sperm. Later they become female, losing the ability to make sperm but acquiring the ability to make eggs, which they can fertilise with the stored sperm.
This biological definition of sex has been swept up into debates over the status of transgender people in society. Some philosophers and gender theorists define a ‘woman’ as a biologically female human being. Others strongly disagree. I’m addressing those who reject the very idea that there are two biological sexes. Instead, they argue, there are many biological sexes, or a continuum of biological sexes.
There’s no need to reject how biologists define the sexes to defend the view that trans women are women. When we look across the diversity of life, sex takes stranger forms than anyone has dreamt of for humans. The biological definition of sex takes all this in its stride. It does so despite the fact that there are no more than two biological sexes in any species you’re likely to have heard of. To many people, that might seem to have ‘conservative’ implications, or to fly in the face of the diversity we see in actual human beings. I will make clear why it does not.
I call this the ‘biological’ definition of sex because it’s the one biologists use when studying sex – that is, the process by which organisms use their DNA to make offspring. Many philosophers and gender theorists will protest at making the creation of offspring foundational to how we define sex or distinguish different sexes. They’re surely right that sex as a social phenomenon is much richer than that. But the use of DNA to make offspring is a central topic in biology, and understanding and explaining the diversity of reproductive systems is an important scientific task. Gender theorists are understandably worried about how the biology of sex will be applied – or misapplied – to humans. What they might not appreciate is why biologists use this definition when classifying the mind-stretching forms of reproduction observed in limpets, worms, fish, lizards, voles and other organisms – and they might not understand the difficulties that arise if you try to use another definition.
Many people assume that if there are only two sexes, that means everyone must fall into one of them. But the biological definition of sex doesn’t imply that at all. As well as simultaneous hermaphrodites, which are both male and female, sequential hermaphrodites are first one sex and then the other. There are also individual organisms that are neither male nor female. The biological definition of sex is not based on an essential quality that every organism is born with, but on two distinct strategies that organisms use to propagate their genes. They are not born with the ability to use these strategies – they acquire that ability as they grow up, a process which produces endless variation between individuals. The biology of sex tries to classify and explain these many systems for combining DNA to make new organisms. That can be done without assigning every individual to a sex, and we will see that trying to do so quickly leads to asking questions that have no biological meaning.
While the biological definition of sex is needed to understand the diversity of life, that doesn’t mean it’s the best definition for ensuring fair competition in sport or adequate access to healthcare. We can’t expect sporting codes, medical systems and family law to adopt a definition simply because biologists find it useful. Conversely, most institutional definitions of sex break down immediately in biology, because other species contradict human assumptions about sex. The United States’ National Institutes of Health (NIH) uses a chromosomal definition of sex – XY for males and XX for females. Many reptiles, such as the terrifying saltwater crocodiles of northern Australia, don’t have any sex chromosomes, but a male saltie has no trouble telling if the crocodile that has entered his territory is a male. Even among mammals, at least five species are known that don’t have male sex chromosomes, but they develop into males just fine. Gender theorists have extensively criticised the chromosomal definition of human sexes. But however well or badly that definition works for humans, it’s an abject failure when you look at sex across the diversity of life.
The same is true of ‘phenotypic sex’, the familiar idea that sex is defined by the typical physical characteristics (phenotypes) of males and females. Obviously, this approach will produce completely different definitions of male and female for humans, for worms, for trees and so forth. Incubating eggs inside your body, for example, is a female characteristic in humans but a male one in seahorses. That doesn’t mean that human institutions can’t use the phenotypic definition. But it isn’t useful when studying the common patterns in the genetics, evolution and so forth of female humans, female seahorses and female worms.
Understanding the complex ways in which chromosomes and phenotypes relate to biological sex will make clear why the biological definition of sex shouldn’t be the battleground for philosophers and gender theorists who disagree about the definition of ‘woman’. There might be very good reasons not to define ‘woman’ in this way, but not because the definition itself is poor biology.
Why did sexes evolve in the first place? Biologists define sex as a step towards answering this question. Not all species have biological sexes, and biology seeks to explain why some do and others don’t. The fact that no species has evolved more than two biological sexes is also a puzzle. It would be quite straightforward to engineer a species that has three, but none has evolved naturally.
Many species reproduce asexually, with each individual using its own DNA to create offspring. But other species, including our own, combine DNA from more than one organism. That’s sexual reproduction, where two sex cells – gametes – merge to make a new individual. In some species, these two gametes are identical; many species of yeast, for example, make new individuals from two, identical gametes. They reproduce sexually, but they have no sexes, or, if you prefer, they have only one sex. But in species that make two different kinds of gamete – and where one gamete of each kind is needed to make a new organism – there are two sexes. Each sex makes one of the two kinds of gamete.
In complex multicellular organisms, such as plants and animals, these two kinds of gamete are very different. One is a large, complex cell, what we’d typically call an ‘egg’. It’s similar to the eggs produced by asexual species, which can develop into a new organism all on their own. Many species of insect and some lizards, snakes and sharks can reproduce using just an egg cell. The other kind of gamete is a much smaller cell that contains very little beyond some DNA and some machinery to get that DNA to the larger gamete. We are familiar with these two kinds of gametes from human eggs and sperm.
There’s no obvious reason why complex multicellular organisms need to have two kinds of gamete, or why these two kinds are so different in size and structure. It’s perfectly possible to make three or more different kinds of gamete, or gametes that vary continuously, just as people vary continuously in height. One question that biologists seek to answer, then, is why those forms of sexual reproduction aren’t observed in complex organisms such as animals and plants.
Earthworms are hermaphrodites: one part of the worm produces sperm and another part produces eggs
When a species produces two different kinds of gamete, biologists call this ‘anisogamy’, meaning ‘not-equal-gametes’. Some anisogamic species have separate sexes, like humans do, where each individual can produce only one kind of gamete. Other anisogamic species are hermaphrodites, where each individual produces both kinds of gamete. Because they produce two kinds of gametes, hermaphroditic species still have two biological sexes – they simply combine them in one organism. When a biologist tells you that earthworms are hermaphrodites, they mean that one part of the worm produces sperm and another part produces eggs.
Some single-celled and very simple multicellular organisms have evolved something called ‘mating types’. These are gametes that are identical in size and structure, but in which the genome of each gamete contains genetic markers that affect which other gametes it can combine with. Typically, gametes with the same genetic marker can’t recombine with one another. Some species have many hundreds of these ‘mating types’, and newspapers often report research into this phenomenon under headlines such as: ‘Scientists discover species with hundreds of sexes!’ But, formally, biologists refer to these as ‘mating types’, and reserve the term ‘sexes’ for gametes that are different in size and structure.
Why distinguish between these two phenomena? One reason is that the evolution of anisogamy – gametes that differ in size and structure – explains the later evolution of sex chromosomes, sex-associated physical characteristics and much more. But the existence of mating types doesn’t have these dramatic knock-on evolutionary effects. Another reason to keep the distinction is that anisogamy and mating types are thought to have evolved via different evolutionary processes. One theory is that anisogamy appeared when mating-type genome markers somehow became linked to genes that controlled the size of the gamete, or mutated in some way that affected gamete size. These differences in gamete size would then kickstart the evolution of sexes.
The evolution of sex seems to be strongly associated with multicellularity, so the obvious place to look for a shift from mating types to sexes is in organisms that sit at the multicellular boundary – such as algae, which sometimes exist as single-celled organisms, and sometimes as colonies of cells. And indeed, there are species of algae where gametes are just a little bit anisogamous, blurring the distinction between mating types and sexes. There’s much we don’t know about how sex evolved, and how it might have evolved differently across species. But the point is that sexes and mating types are very different phenomena, with different causes and consequences.
The fact that sex evolved in some species but not others tells us something important about how biologists think about sex. Many cultures take the difference between male and female to be something fundamental, and label other natural phenomena such as the Sun and the Moon as male or female. But for biologists, the separation between male and female is no more fundamental or universal than photosynthesis or being warm-blooded. Some species have evolved these things, and some haven’t. They exist when they do only because of the local advantages they afforded in evolutionary competition.
So why did some species evolve two, distinct sexes? To answer this question, we need to forget about creatures with complex sex organs and mating behaviours. These evolved later. Instead, think of an organism that releases its gametes into the sea, such as coral, or into the air, such as fungal spores. Next, consider that there are two goals that any gamete must achieve if it’s to reproduce sexually. The first is finding and recombining with another gamete. The second is producing a new individual with enough resources to survive. One widely accepted idea, then, is that the evolution of sexes reflects a trade-off between these goals. Because no organism has infinite resources, organisms can either produce many small gametes, making it more likely that some of them will find a partner, or produce fewer but larger gametes, making it more likely that the resulting individual will have what it needs to survive and thrive.
Since the 1970s, this idea has been used to model how anisogamic species might have evolved from species with only one kind of gamete. As mutations introduce differences in gamete size, two winning strategies emerge. One is to produce a large number of small gametes – too small to create viable offspring unless they recombine with a larger, well-provisioned gamete. The other winning strategy is to produce a few, large, well-resourced gametes that can create viable offspring, no matter how small the recombinant they end up merging with. Intermediate approaches, such as producing a moderate number of moderately well-provisioned gametes, don’t do well. Organisms that try to follow the ‘middle way’ end up with gametes less likely to find a partner than smaller gametes, and more likely to have insufficient resources than larger gametes. When the two successful complementary strategies have evolved, fresh evolutionary pressures make the gametes even more distinct from one another. For example, it can be advantageous for the small gametes to become more mobile, or for the large, immobile gametes to send signals to the mobile ones.
Once anisogamy has evolved, it shapes many other aspects of reproductive biology. Most species of limpet – shellfish that you see on rocks at the beach – are sequential hermaphrodites. When young and small they are male, and when mature and large they become female. This is believed to be because small limpets don’t have sufficient resources to produce large female gametes, but they’re capable of producing the smaller male ones. In some other species, successful males can arrest their growth and remain small (and male) for their entire life.
Chromosomes aren’t ‘male’ or ‘female’ because these bits of DNA define biological sex. It’s the other way around
Sequential hermaphroditism occurs in the opposite direction too. Australian snorkellers love to spot the large blue males of the eastern blue groper, but it’s rare to see more than one. Most groper are smaller, brown females. They are all born female and become sexually mature after a few years, when 20 or 30 cm in length. At around 50 cm, they change sex and acquire other male characteristics, such as being blue. Unlike the limpet, the main problem facing a male groper is controlling a territory on the reef, so becoming male when you’re small is a losing strategy.
Biology aims to understand the extraordinary diversity of ways in which organisms reproduce themselves, as well as to identify common patterns, and to explain why they occur. In general, organisms become sexually mature when they reach an optimal size for reproduction. This optimal size is often different for the two sexes, because the two sexes represent divergent strategies for reproduction. The limpet and the groper are two of many examples. In constructing these explanations, biological sex is defined as the production of one type of viable anisogamous gamete. If we defined sex in some other way, it would be hard to see the common patterns across the diversity of life, and hard to explain them.
So-called ‘sex chromosomes’, such as the XX and XY chromosome pairs seen in humans, are often brandished as something that’s fundamental to sex. It’s partly the inadequacy of this definition that drives scepticism about the existence of two, discrete biological sexes. ‘Molecular genetics is likely to require a shift from binary sex to quantum sex, with a dozen or more genes each conferring a small percentage likelihood of male or female sex that is still further dependent on micro- and macroenvironmental interactions,’ writes the gender scholar Vernon Rosario.
But any biologist already knows that there’s more to sex determination than chromosomes and genes, and that male and female sex chromosomes are neither necessary nor sufficient to make organisms male and female. Several species of mammal, all rodents of one kind or another, have completely lost the ‘male’ Y chromosome, but these rats and voles all produce perfectly normal, fertile males. Other groups of species, such as crocodiles and many fish, have neither sex chromosomes nor any other genes that determine sex. Yet they still have two, discrete biological sexes. The Australian saltwater crocodile, whom we met before, lays eggs that are very likely to develop into gigantic, highly territorial males if incubated between 30 and 33 degrees Celsius. At other temperatures, genetically identical eggs develop into much smaller females.
The reality is that chromosomes aren’t called ‘male’ or ‘female’ because these bits of DNA define biological sex. It’s the other way around – in some species that reproduce using two discrete sexes, those sexes are associated with different bits of DNA. But in other species this association is either absent or unreliable. Medical institutions use a chromosomal definition of sex because they judge, rightly or wrongly, that this is a reliable way of categorising humans. But humans really aren’t the best place to start when trying to understand sex across the diversity of life.
So much for genes. But perhaps sex could be defined by the physical characteristics that organisms develop, which then ‘add up’ to constitute an organism’s sex? An organism with more female than male characteristics would be more female than male and vice-versa. That’s a reasonable way to think about sex, and this idea of ‘phenotypic sex’ is widely used. But if we apply the biological definition of sex, some of the individuals who are ‘in the middle’ as far as sex-associated characteristics go are bona fide members of one biological sex. Others are not clearly members of either biological sex.
Nothing in the biological definition of sex requires that every organism be a member of one sex or the other. That might seem surprising, but it follows naturally from defining each sex by the ability to do one thing: to make eggs or to make sperm. Some organisms can do both, while some can’t do either. Consider the sex-switching species described above: what sex are they when they’re halfway through switching? What sex are they if something goes wrong, perhaps due to hormone-mimicking chemicals from decaying plastic waste? Once we see the development of sex as a process – and one that can be disrupted – it is inevitable that there will be many individual organisms that aren’t clearly of either sex. But that doesn’t mean that there are many biological sexes, or that biological sex is a continuum. There remain just two, distinct ways in which organisms contribute genetic material to their offspring.
What’s more, the physical characteristics of an organism can be labelled as ‘male’ or ‘female’ only if there is already a definition of sex. What’s so ‘male’ about a groper being blue as opposed to brown? Many male organisms are brown. What’s so ‘female’ about incubating eggs in a womb? After all, in many pipefish and seahorse species the male incubates the eggs in his brood pouch. What makes this part of the hermaphroditic earthworm male and that part female? Gender studies scholars have noticed this logical discrepancy, and some have gone on to argue that the sexes must therefore be defined in terms of gender. But from a biological perspective, what makes an observable physical characteristic ‘male’ or ‘female’ is not its association with gender, but its association with something more tangible: the production of one or other of the two kinds of gamete.
This explains why the existence of individuals with combinations of male and female characteristics doesn’t show that biological sex is a continuum. These organisms have a combination of characteristics associated with one biological sex and characteristics associated with the other biological sex. They do not have some part of the ability to make small gametes combined with some part of the ability to make large gametes. Their phenotypic sex might be intermediate, but their biological sex is not. Being fully biologically male and fully biologically female – hermaphroditic – can be an effective evolutionary strategy, and we have encountered several hermaphroditic species already. But making both kinds of gametes incompletely would be an evolutionary dead-end.
Like phenotypic characteristics, sex chromosomes can be more or less reliably associated with biological sex. The eastern three-lined skink, an Australian lizard, has sex chromosomes, and under some circumstances XY skinks become male and XX skinks become female, just as in humans. But in cold nests, every skink becomes male, whatever their chromosomes. By ‘becomes male’, biologists mean that they grow up to produce small gametes – sperm.
No animal is conceived with the ability to make sperm or eggs (or both). This ability has to grow
This effect of temperature on sex is not surprising, as many reptile species produce genetically identical offspring whose sex is determined by incubation temperature. What’s more surprising is that varying the size of the egg yolk in this species of skink can produce both sexes with the ‘wrong’ sex chromosomes: XX males and XY females. The skink seems to have three mechanisms for determining sex – chromosomes, temperature and hormones in the yolk. This is not a mere quirk of nature. The skink is one of many species that actively control the sex of their offspring, responding to environmental cues that predict whether male or female offspring have better chances of surviving and reproducing.
If all species were like the skink, we probably wouldn’t label sex chromosomes as ‘male’ or ‘female’. After all, we don’t think of extreme nest temperatures as ‘female’ and intermediate temperatures as ‘male’, merely because they produce male and female crocodiles or male and female geckos. We think of sex chromosomes as male or female because we focus on species where they are reliably associated with the production of male or female gametes.
Sex chromosomes play much the same role in sex determination as nest temperatures and hormones. They’re simply mechanisms that organisms use to turn genes on and off in offspring so that they develop a biological sex. No animal is conceived with the ability to make sperm or eggs (or both). This ability has to grow, through a cascade of interactions between genes and environments. In some species, once an individual acquires a sex, it remains that sex for the rest of its life. In others, individuals can switch sex one or more times. But in every case, the underlying mechanisms are designed to grow organisms that make either male or female gametes (or both). The other changes the body undergoes as it becomes male, female or hermaphroditic are designed to fit the reproductive strategies that this species has evolved.
These mechanisms by which organisms develop or switch biological sex are complex, and many factors can interfere with them. So they produce a lot of phenotypic diversity. Sometimes, organisms grow up able to make fertile gametes, but otherwise atypical for their biological sex. Sometimes, they grow up unable to make fertile gametes of either kind. This is usually an accident, but sometimes by design. In bees, eggs that aren’t fertilised develop into males, so male bees have half as many chromosomes as female bees. Meanwhile, all fertilised eggs start to develop into females, but most of them never complete their sexual development. The queen sends chemical signals that block the development of the worker bee’s ovaries at an early stage. So worker bees are ‘female’ in the extended sense that they would develop into fertile females if they weren’t actively prevented from doing so. Occasionally, worker bees manage to evade these controls and lay their own eggs. They are not popular with beekeepers, who select against these mutant strains.
The diversity of outcomes in individual sexual development doesn’t mean that there are many biological sexes or that biological sex is a continuum. Whatever the merits of those views for chromosomal sex or phenotypic sex, they are not true of biological sex. A good way to grasp this is to imagine a species that really does have three biological sexes. Biotechnologists have proposed curing mitochondrial diseases by removing the nucleus from an egg with healthy mitochondrial DNA, and inserting a new nucleus containing the nuclear DNA from an unhealthy egg and the nuclear DNA from a sperm. The resulting child would have three genetic parents.
Now imagine if there was a whole species like this, where three different kinds of gametes combined to make a new individual – a sperm, an egg and a third, mitochondrial gamete. This species would have three biological sexes. Something like this has actually been observed in slime moulds, an amoeba that can, but need not, get its mitochondria from a third ‘parent’. The novelist Kurt Vonnegut imagined an even more complex system in Slaughterhouse-Five (1969): ‘There were five sexes on Tralfamadore, each of them performing a step necessary in the creation of a new individual.’ But the first question a biologist would ask is: why haven’t these organisms been replaced by mutants that dispense with some of the sexes? Having even two sexes imposes many extra costs – the simplest is just finding a mate – and these costs increase as the number of sexes required for mating rises. Mutants with fewer sexes would leave more offspring and would rapidly replace the existing Tralfamadorians. Something like this likely explains why two-sex systems predominate on Earth.
We can also imagine a species where biological sex really does form a continuum. Recall that some algae have slightly anisogamous gametes, much closer together than sperm and eggs. We can imagine a more complex organism using this system, with some slightly smaller gametes and some slightly larger ones. Successful reproduction might require two gametes that, when added together, are big enough but not too big. But the sexually reproducing plants and animals that actually exist all have just two, very different kinds of gamete – male and female. They’re not merely different in size, they’re fundamentally different in structure. This is the result of competition between organisms to leave the greatest number of genetic descendants. In complex multicellular organisms such as plants and animals, we know of only three successful reproductive strategies: two biological sexes in different individuals, two biological sexes combined in hermaphroditic individuals, and asexual reproduction. Some species use one of these strategies, some use more than one.
Human beings have come up with many ways to classify the diversity of individual outcomes from human sexual development. People who want to apply the biological definition of sex to humans should recognise that it’s ill-suited to do what many human institutions want, which is to sort every individual into one category or another. What sex are worker bees? They are sterile workers whose genome was designed by natural selection to terminate ovary development on receipt of a signal from the queen bee. They share much of the biology of fertile female bees – but if someone wants to know ‘Are worker bees really female?’, they’re asking a question that biology simply can’t answer.
Nor is being a sterile worker a third biological sex alongside male and female. This is easier to see in ants, where there is more than one sterile caste. Workers, soldiers, queens and male ‘flying ants’ each have specialised bodies and behaviour, but there are not four biological sexes of ant. Workers and soldiers are both ‘female’ in an extended sense, but not in the full-blown sense that queen ants are female. There is a human imperative to give everything a sex, as mentioned above, but biology doesn’t share it.
The biological definition of sex wasn’t designed to ensure fair sporting competition, or settle healthcare disputes
Juvenile organisms and postmenopausal human females also can’t produce either kind of gamete. Juveniles are assigned to the sex they have started to grow into. But once again, this is more complicated than it seems when we focus only on humans. In almost all mammals, sexual differentiation is initiated by a region of the Y chromosome, so a mammalian egg can become either male or female. In birds, it’s the other way around – the egg carries the sex-determining W chromosome, so sperm can become either male or female. After fertilisation, therefore, we can say that an individual mammal or bird has a sex in the sense that it has started to grow the ability to produce either male or female gametes. With a crocodile or a turtle, though, we’d have to wait until nest temperature had its sex-determining effect. But that doesn’t mean that we need to create a third biological sex for crocodile eggs!
More importantly, nothing guarantees that any of these organisms, including those with sex chromosomes, will continue to grow to the point where they can actually produce male or female gametes. Any number of things can interfere. From a biological point of view, there is nothing mysterious about the fact that organisms have to grow into a biological sex, that it takes them a while to get there, and that some individuals develop in unusual or idiosyncratic ways. This is a problem only if a definition of sex must sort every individual organism into one sex or another. Biology doesn’t need to do that.
In human populations, there are plenty of individuals whose sex is hard to determine. Biologists aren’t blind to this. The definition of biological sex is designed to classify the human reproductive system and all the others in a way that helps us to understand and explain the diversity of life. It’s not designed to exhaustively classify every human being, or every living thing. Trying to do so quickly leads to questions that have no biological meaning.
Human societies can’t delegate to biology the job of defining sex as a social institution. The biological definition of sex wasn’t designed to ensure fair sporting competition, or to settle disputes about access to healthcare. Theorists who want to use the biological definition of sex in those ways need to show that it will do a good job at the Olympics or in Medicare. The fact that it’s needed in biology isn’t good enough. On the other hand, whatever its shortcomings as an institutional definition, the concept of biological sex remains essential to understand the diversity of life. It shouldn’t be discarded or distorted because of arguments about its use in law, sport or medicine. That would be a tragic mistake.
The author’s research is supported by the Australian Research Council and the John Templeton Foundation. He would also like to thank Nicole Vincent, Jussi Lehtonen, Stefan Gawronski and Joshua Christie for their feedback on earlier drafts.