If you split yourself down the middle to become two people, would you survive the process? And, if you did, would your other half be your child, your clone or your sibling? Would this create two instances of the same you, existing simultaneously in two places at the same time; or would it create two entirely new people, causing you to suddenly cease to exist? While such thought experiments raise baffling questions about personal identity, there is a more fundamental problem I want to consider: would splitting in two be an instance of reproduction or an entirely different kind of process?
When we think about how organisms reproduce, we don’t tend to think of splitting bodies. We think of sex. We tend to think of animals such as panda bears, leopards, ravens or any other large multicellular organisms having sex, becoming pregnant (or laying fertilised eggs), and giving birth. It isn’t surprising that this is how we think new organisms come into existence. Sexual reproduction is, after all, the form of reproduction that nature has selected for creatures like us. But sex is not the way most reproduction takes places.
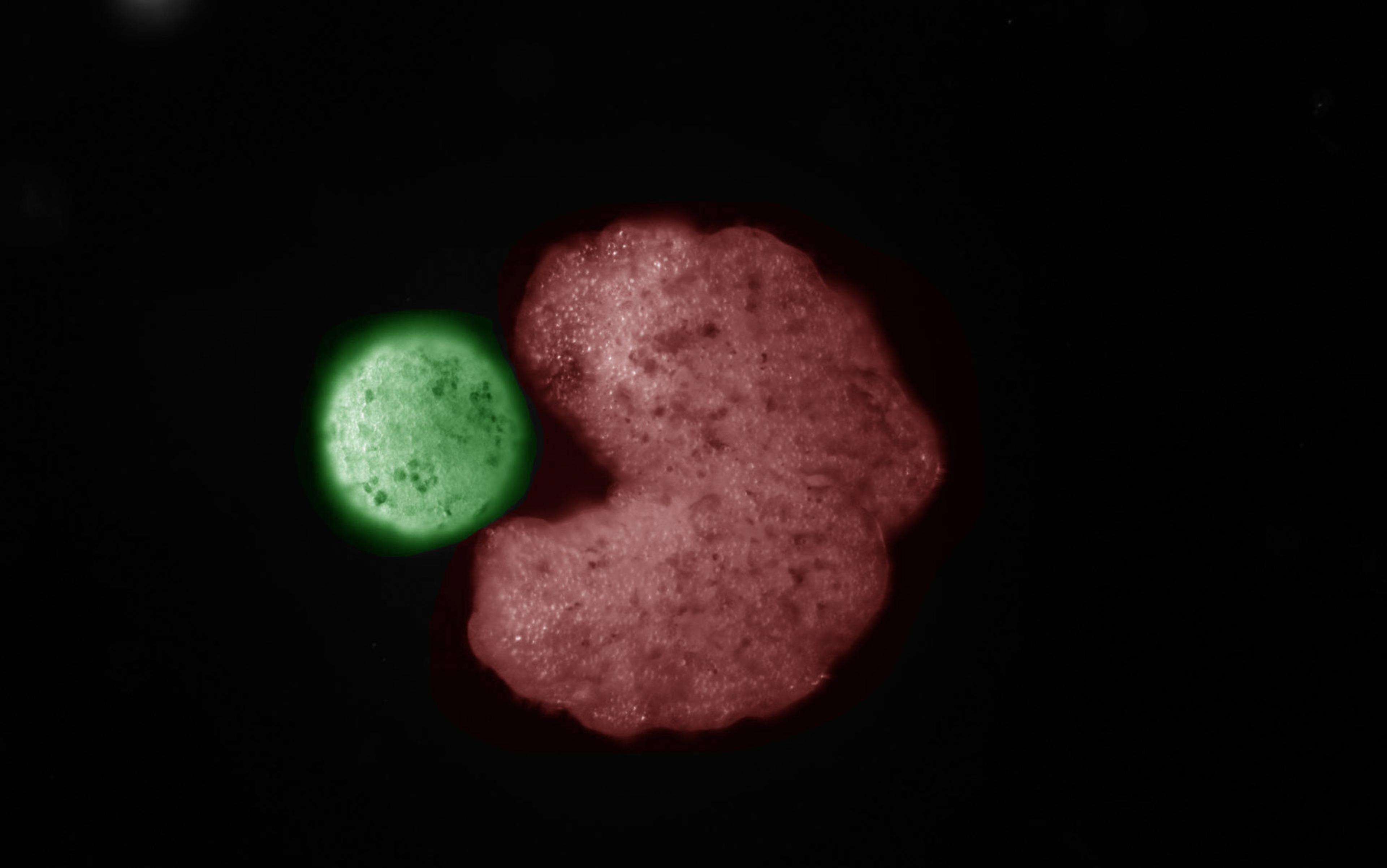
Most forms of life on this planet create other living beings through asexual processes – and there are many ways this can happen (as we’ll see). Some of the most common forms are similar to the thought experiment above: a body splits in two. Nearly all prokaryotic microbes, such as bacteria, reproduce through various forms of this process, such as binary fission (when a body separates into two new bodies). However, it’s not always clear what kinds of relation result from fission, as in the thought experiment above.
As a philosopher of biology, some of my research considers what counts as a biological individual, and how biological individuals are connected to each other. It seems to me that our concept of reproduction often distorts how we think about organisms coming into existence. This isn’t surprising: it’s a concept developed by 18th-century naturalists who knew very little about the fission processes we can observe today. What needs to be done to align our concept of biological reproduction with the fission processes seen throughout so much of life? Where do we start?
To think about this, we first need to figure out how we understand reproduction itself. This may seem like a theoretical exercise, but a lot is at stake in how we define the concept. Our ability to identify species, aliens and even the branches on the tree of life hinges on the concept.
Composed of reproductive links between organisms, much of the tree of life rests on our understanding of reproduction. How we think about vertical lines of descent (between parents and offspring across generations) can have massive implications for our evolutionary models. Say an organism survives reproductive fission and has 40 offspring. All those offspring – 40 bodies that split off over time – are one generation away from their parent. However, if you think an organism doesn’t survive fission, then the organisms born after 40 splits are 40 generations removed from the first organism to break into two. Thus, whether two organisms are understood as being just a few generations apart or (potentially) millions of generations apart hinges on how you understand reproductive fission.
Astrobiologists are also heavily invested in how reproduction is defined. If we’re ever going to find life outside Earth, they will need to know how to tell the living from the non-living. One of the more prominent theories about how to do this is by determining whether an extraterrestrial candidate can reproduce or pass on its genes. According to this theory, a capacity to reproduce could be one of the defining characteristics of life. Reproduction is also central to the way we define species. The German-born evolutionary biologist Ernst Mayr ’s biological species concept – the most common concept used to define a species – treats them as interbreeding (or potentially interbreeding) populations. Updating how we think about reproduction, then, creates a series of difficult identity problems that challenge the foundations of many of our ideas about life.
It’s perhaps surprising then that the concept of biological reproduction, which underpins our ability to define species, the search for alien life and the concept of evolution, emerged only around the mid-18th century. The work of the French naturalist Georges-Louis Leclerc, Comte de Buffon and the French philosopher Pierre-Louis Maupertuis marks the beginning of the idea that we think of as ‘reproduction’ today. Prior to their work, organisms were believed to form through generation, not reproduction. The French biologist François Jacob’s The Logic of Life (1970) nicely illustrates the importance of the historical shift between these concepts.
From the perspective of generation, a human child with a frog’s face seems perfectly reasonable
Before reproduction, it was generally believed that animals were generated in a similar way to anything else (houses or statues, for example) through the right combination of matter and form. Thinkers as far back as Plato and Aristotle believed that life was created by an immutable set of forms or patterns that could recombine in different materials or, as Jacob says, through different combinations of ‘visible structures’. Forms were supposed to function, one might say, like cookie cutters. Thinking about living organisms in this way rendered them similar to other physical objects: just as the bronze of a statue could take on different forms, so could life. The only thing that set life apart from everything else was that it needed an extra nudge to get going. Some believed this nudge was divine; for others, a causal impetus was contained in the seed or germ; or it was inherent in the final cause of the form in the way that a statue comes into being by taking the right kind of shape.
This is why talk of ‘monsters’ abounded prior to the 18th century – tales of animals with deformities, or creatures with amalgamations of different animal parts. In the 16th century, the French surgeon Ambroise Paré dedicated an entire book to them called On Monsters and Marvels where he reports on all sorts of implausible chimeras, such as a human child with a frog’s face. Paré wasn’t the first. Millennia earlier, Aristotle gave an extensive account of ‘monsters’ in his On the Generation of Animals as well. From Aristotle to Paré and beyond, talk of animals possessing strange mixes of parts seemed reasonable because the concept of generation isn’t constrained by the past – each instance of generation could recombine form and matter in new ways. The idea behind this is somewhat like building with Lego blocks. The blocks connect in many ways, and are limited only by their own shapes and how they can link together. Similarly, the forms animals could take were constrained only by the kinds of matter that made them up – flesh, blood, bone, feathers, scales – and the principles that governed all physical objects. From the perspective of generation, a human child with a frog’s face seems perfectly reasonable.
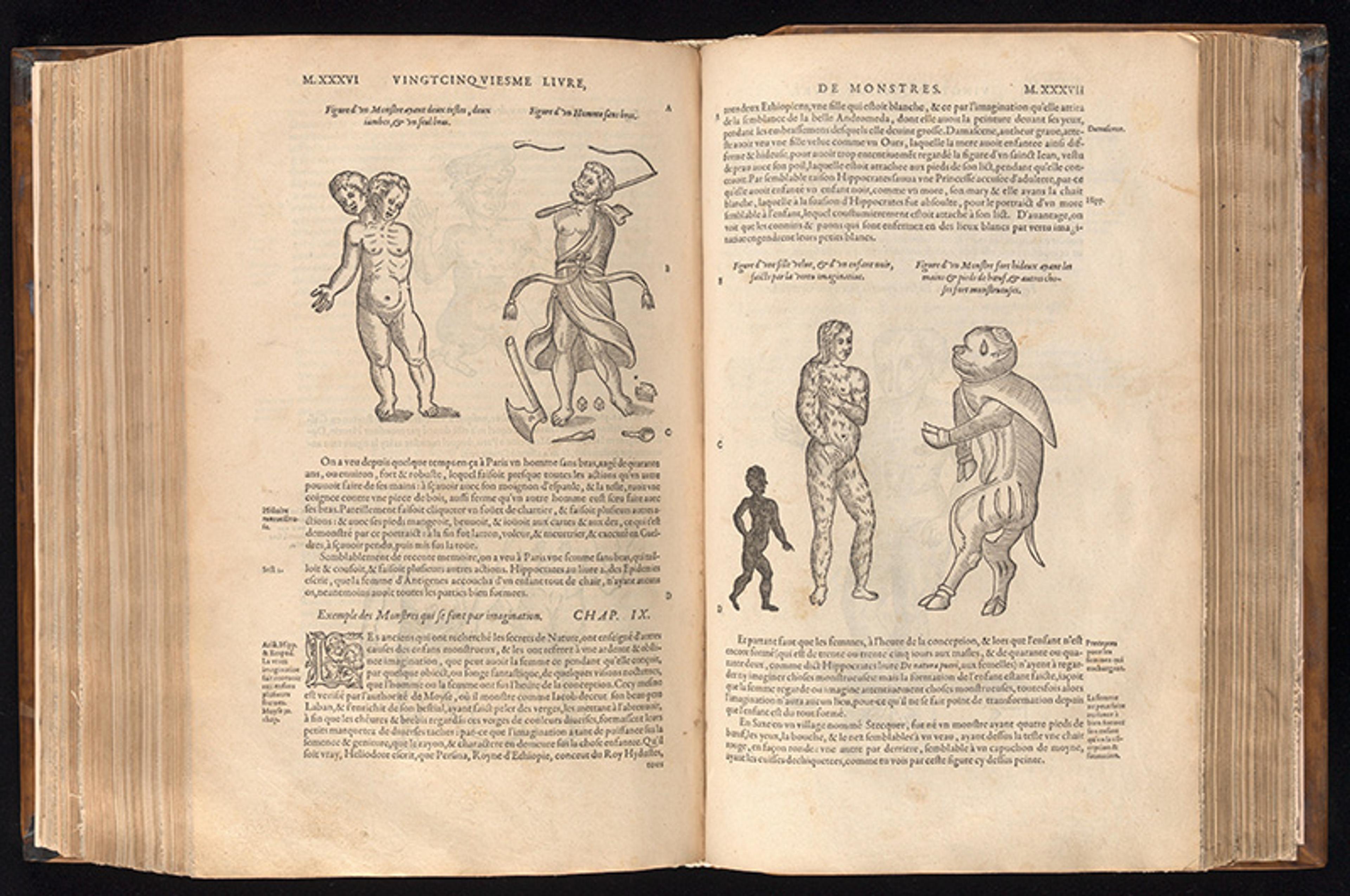
From Monsters and Marvels by Ambroise Paré. Courtesy the Wellcome Collection
Contrast this with the modern notion of reproduction. From today’s vantage point, understanding an organism requires understanding its evolutionary history, the lineage of ancestors from which it’s descended. Naturalists in the 18th century began to notice the importance of biological history when they started experimenting with hybrids by breeding different species. When testing which animals could hybridise, they observed that it was rare for animals belonging to different species to successfully interbreed, and, even when they did, very few of the resulting hybrids were fertile. This made the idea of ‘monsters’ created through generation look quite implausible, if not impossible. Armed with such observations, naturalists and philosophers came to see that generation wasn’t good at explaining how new organisms came into existence.
Organisms weren’t like houses or statues – they weren’t ‘generated’ in the same way a builder makes a house with matter (wood) and form (a blueprint). Instead, an organism’s anatomy and physiology were constrained by what they’d inherited from their ancestors. Understanding a specific animal required understanding its history or lineage. A house or statue can be understood by its physical structure but, to really understand life, 18th-century naturalists realised they needed to go beyond the physical structure of the organism. In this way, the difference between generation and reproduction involves distinctions between space and time: generation was concerned with space (form, physical structure), but reproduction introduced time (a historical component). The conceptual shift from generation to reproduction marks one of the most important transitions in the history of biology. It is one of the factors that led to Charles Darwin’s epoch-making theory of evolution.
Of course, since the 18th century, biologists have learned vastly more than just this about life. The biological philosopher Maureen O’Malley has shown that, by most measures, the vast majority of life on Earth is microbial. Contained within about one tonne of soil are 1016 prokaryotic organisms, significantly more than the ‘mere’ 1011 stars in the Milky Way. Conservative estimates suggest that half of Earth’s total biomass is made up of these tiny prokaryotic microbes. In Earth’s oceans, they constitute closer to 90 per cent of the biomass.
In terms of the number of species, microbes put the rest of the planet’s life to shame. Depending on whom you talk to, there are at least a trillion different species of microbes on Earth (yes, a trillion species, not microbes). It is not an exaggeration to say that life on Earth is microbial, with the comparatively rare exception of the odd macro-organism. How are these microbes relevant to our concept of reproduction? Their staggering presence means that life as we know it is overwhelmingly, by many orders of magnitude, asexual. And this extends far beyond the world of microbes. There are a multitude of plant species that reproduce asexually, or through a combination of both sexual and asexual processes. Perhaps you’ve owned one of these asexual plants? A shoot cut from the common houseplant Epipremnum aureum, sometimes known as ‘Devil’s Ivy’, will continue to grow when replanted in another pot. That’s a form of asexual reproduction. Some animals – aphids, flatworms and even certain lizards – reproduce through other kinds of asexual processes, too. So, aside from bringing new organisms into existence, what do these processes have in common with sexual reproduction?
Currently, perhaps the best candidate for a general account of reproduction is offered by the American philosopher James Griesemer. He suggests that reproductive processes generally share two features: first, there must be an overlap of material parts (ie, the physical material that makes up an organism) between parents and offspring; and, second, those parts must go on to develop. And while there are other accounts of reproduction, Griesemer presents a compelling case. His first criterion, material overlap, requires that some material part of an offspring has been some part of the parent. This criterion is important because, without it, all sorts of processes that are obviously not instances of biological reproduction might qualify as reproduction. For example, a photocopier making copies of a sheet of paper is not an instance of reproduction because no material from the original paper is transferred to the copy. That’s why photocopying is mere production rather than reproduction.
The second criterion, development, is also important to block other cases of production that are obviously not instances of reproduction either. For example, cutting a piece of paper in half produces two sheets of paper from one original. In this case, part of the material from the original is used to produce a new sheet of paper, but we don’t want to call this reproduction either. For this reason, the criterion of development is necessary. For a process to be one of genuine reproduction, not only must material transfer from parent to offspring, but the offspring must be able to develop the capacity to reproduce, too. The sheet of paper fails this test. Thus, Griesemer offers us some good reasons to think that what’s happening with sexual reproduction and asexual reproduction shares important similarities that can’t be disregarded – similarities that make these processes quite unlike the production of houses or statues.
Perhaps fission is not only reproduction but also a very strange kind of death
Now let’s think about this account of reproduction in terms of our strange case of fission or splitting. After all, fission-like processes are the way most forms of life reproduce. What happens when a microbe, such as an E coli bacterium, undergoes binary fission? Much like the sheet of paper that’s cut in half, during binary fission a single bacterium splits into two bacteria. And when an E coli bacterium splits, the two resulting bacteria are almost perfectly identical to one another. Reproduction through these kinds of fission processes meets Griesemer’s two criterions as there is both material overlap between parents and offspring, and the two bacteria that emerge from a split bacterium can develop the capacity to have offspring of their own.
But how do we think about the identity of these two bacteria? It’s similar to the thought experiment of a person splitting down the middle to become two people. It’s the stuff of science fiction, not to mention the starting point of philosophical debates about the nature of personal identity. Take, for example, the American philosopher David Lewis who has argued that if someone split to become two other people, both would be instances of that single person because one person possesses a multitude of identities (some of which could splinter off in the future). In arguing for this, he’s not suggesting that we’re all affected by something like multiple personality disorder. Rather, he’s saying that it’s quite common for one thing to possess multiple identities. Think of a street that forks. Imagine you’re driving down a street that is both ‘Route 23’ and ‘Washington Ave’. Then you reach the fork, the street splits, and you follow Route 23. Prior to the fork, the one street had two identities.
But not everyone agrees with Lewis. The British philosopher Derek Parfit argues that one person splitting into two people results in that person’s identity ceasing to exist, in something that seems very unlike biological death, but where it’s not entirely clear if the original person survives. Parfit thinks that each person, after the split, might retain the same beliefs, desires and goals: however, seeing as plants and microbes don’t obviously have these, it’s unclear what of them survives.
These curious identity questions don’t yet have obvious answers, and it’s unclear what kind of ancestral relation fission creates. A lot would likely depend on the particulars of the fission involving ageing, growth and, in particular, symmetry. If there’s perfect symmetry during fission, then perhaps Parfit is correct and certain organisms pop out of existence – meaning fission is not only reproduction but also a very strange kind of death. However, if the fission isn’t symmetrical, then some organisms might be more like Lewis’s streets. If this is the case, perhaps colonies of bacteria are composed of many individuals that share a single identity. It’s clear that these kinds of fission meet Griesemer’s criteria for reproduction. But the problem is that fission doesn’t create clear links from parent to offspring: a clear, vertical lineage of descent. This creates difficulties for our notions of reproduction. To return to the questions that opened this essay, how do we define the relationships between organisms created through fission? Does this process produce clones, siblings or just two instances of the original? To see why, let’s return to the Devil’s Ivy.
Say someone cuts a shoot from a mature Devil’s Ivy plant and replants the shoot in another pot, where it develops into a mature plant. According to Griesemer’s criterion, this counts as an instance of reproduction. But has the plant really reproduced in the sense of 18th-century-style reproduction? Is the original plant really a parent and the shoot its offspring? It’s far from obvious. First, remember that the shoot and the original plant are genetically identical clones. All that changes is physical connectedness, but cutting and replanting a shoot doesn’t resemble something like human birth. Other creatures are also able to persist through physical disconnectedness, like the ‘immortal’ jellyfish Turritopsis dohrnii which, depending on its environmental conditions, moves between being a physically connected mature medusa to an immature stage of disconnected polyps. When food is scarce or it’s otherwise threatened, this jellyfish can break apart into a colony of polyps, and then recombine again when conditions improve (slime moulds do something quite similar). The bodily transformations of Turritopsis dohrnii make the creatures of science fiction look tame.
And then there are cases in the other direction, where two physically disconnected organisms become physically continuous. For example, separate trees (sometimes even different species of trees) can grow together in a process known as inosculation. This happens when the trunks, branches or roots of separate trees come in contact and then, over time, end up grafting to one another. When two physically separate trees grow into one another, do they become a new, singular organism? Philosophers of biology and theoretical biologists are grappling with such questions, dragging the issue of what counts as a biological individual into a metaphysical quagmire to which there are no obvious empirical solutions. Understanding when reproduction has taken place in these cases often depends on intuitions about physical connectedness: how physically connected does an organism need to be to still be an organism? In the case of inosculation, there is no test a biologist could conduct that will tell us whether the original plant is a parent or not. The problem is conceptual.
One possible solution to this conceptual issue can be found in the work of some botanists and biologists from the 1970s (including John Harper and Daniel Janzen) who introduced concepts of ramet and genet to distinguish between the different kinds of individuals that are found among certain plants (and some animals). These concepts were developed to solve the puzzle of how to understand the relationship between genetically identical plants that appeared to be separate individuals, such as dandelions. They introduced the idea that a set of genetically identical individuals form a single, spatially discontinuous ‘genet’. At the same time, each of these individual dandelions, physically separated from the rest, is a ‘ramet’. A group of ramets makes up a genet. This pioneering work shows the very real biological limitations that can come from assumptions about reproduction. Viewing life through the lens of 18th-century naturalists distorts our sense of what an individual looks like because it assumes a world that resembles us. Instead, the concepts of ramet and genet reveal that what counts as an individual can vary, and that parent-offspring relationships are one of many possibilities.
Is each stem of the Devil’s Ivy a possible fork in the road, each with its own identity?
While there are some good reasons to think the ramet and genet solution helps make sense of some curious problems regarding biological individuality, it doesn’t completely solve the problem of how to understand reproduction. It doesn’t tell us how genets reproduce. Griesemer’s criteria for reproduction works for ramets – there is both material overlap and development. Material parts of a ramet are transferred to their ‘offspring’ and those parts go on to develop. But the reproductive process of a ramet (an individual dandelion) is the growth and development of a genet (a disparate grouping of genetically identical dandelions). If that’s right, then how do genets reproduce? From the perspective of a genet, this process looks like development and growth, not reproduction.
Griesemer seems to agree on this front. He acknowledges that there are times when the distinction between reproduction and development collapses, ceasing to be a meaningful division. But it’s important to remember that these cases are not isolated or rare. Processes where the division between reproduction and development become entangled are among the most common biological processes – far more common than the sexual processes we typically think of when we think about reproduction. Just as we’ve discovered that there are very different kinds of biological individuals (eg, ramets and genets), there are also very different kinds of reproductive processes that might not always create parent-offspring relationships. How do we begin to describe these processes? When you cut a shoot from some Devil’s Ivy and replant it elsewhere, has a new entity come into existence or is it more like Lewis’s roads? Is each stem of the Devil’s Ivy a possible fork in the road, each with its own identity, and all contained within a single plant that itself is just one identity of some other plant to which it was once connected?
If you think Lewis is wrong – that a single thing does not contain multiple identities – perhaps what drives your intuition is a sense of spatial connectivity. Intuitively, you might say that this Devil’s Ivy reproduces when a shoot is physically disconnected from its ‘parent’. But is spatial connectedness really the best means of judging when reproduction has occurred? It seems reminiscent of pre-18th-century thinking: an intuition rooted in a very human perspective of the world, which forgets that reproduction has to do with marking history, not only space. So, what can be done to update how we think about biological reproduction?
Rethinking the concepts that we use in biology has led to advances that can be as important as empirical discoveries. Natural philosophers and scientists used to believe that organisms were formed through the process of generation rather than reproduction. They also used to believe that all microbial life was sexual. The concepts of ‘reproduction’ and ‘asexuality’ were introduced in order to help explain what was observed, but that didn’t comport with the conceptual frameworks that were available at the time. These changes led to important advances in how we think about life. It seems we might be at a similar juncture today. Throughout the 20th and 21st centuries, we’ve learned a great deal more about just how much of the biological world is composed of asexual organisms – most of which exhibit various kinds of fission processes.
Unfortunately, this new view of life does not always fit well with the concept of reproduction we inherited from the European naturalists and philosophers of the 18th century. Updating the concept of reproduction might require rethinking whether that plant in your living room or garden, in this moment, is really complete, and fully there? It might be a kind of life that’s very unlike you in that regard. It might be much more like some hard-to-fathom creature from a work of speculative fiction.
Rethinking reproduction could require accepting that asexual life just doesn’t fit our intuitions about identity and individuality – about where one thing begins and another ends. At the very least, it shows that, as life evolved, some of its twists and turns led to distinctly different processes of reproduction and development: but, in other cases, reproduction and development are simply the same process. So, when looking to identify the boundaries of species, the branches on the tree of life, or even what counts as life itself, we should be careful about assumptions that come from our very human perspective of biology. New observations made by biologists not only show us just how amazing and fascinating life is, but these observations also challenge some of the most elemental concepts we use to understand life too. Where one organism ends and another one begins may simply depend on whether you’re thinking about it as an object in space or an object in time.